
Predicting the effect of climate change on the abundance and distribution of tsetse flies
November 30th, 2015
Author: Glyn Vale, co-author: John Hargrove
Tsetse flies (genus Glossina) occur only in sub-Saharan Africa where they infest about 40% of the land surface. They can threaten health and agriculture by transmitting the parasites that cause the potentially fatal diseases of sleeping sickness in humans and nagana in livestock. The fact that only parts of sub-Saharan Africa are infested is attributable to many causes, including the abundance of suitable host animals and vegetation, and the institution of control measures. It is also attributable to some places being too hot or too cold for the flies. This raises the possibility that climate change will affect the abundance and distribution of tsetse - the leading questions being how great and how rapid the effects will be. SACEMA is addressing these questions by a combination of field work and simulation modelling.
Field data
Some of the field work assesses the degree of climate change that has already occurred and gauges the way that this has affected the flies. Other work studies the seasonal abundance and distribution of the tsetse, in the expectation that a fuller understanding of the impact of seasonal weather will allow predictions for longer term drifts in climate. The research is being performed in Tanzania, in the neighbourhood of the Serengeti National Park and in northern Zimbabwe, particularly at and near Rekomitjie Research Station in the Mana Pools National Park of the Zambezi Valley. Records for temperature, rainfall and tsetse abundance at Rekomitjie go back to 1959. The two species of tsetse occurring at Rekomitjie, Glossina morsitans and G. pallidipes, have been intensively studied over almost all of this period and more is known about their behaviour and ecology than for any other tsetse species anywhere.
The reality of climate change is indicated by the fact that the mean daily minimum temperature at Rekomitjie has increased significantly by 1.66oC since 1959. The mean daily maximum has also increased significantly, by 0.92oC, although there has been no significant change in rainfall. Most of the increase in temperature seems to have occurred in the dry season, i.e., June to November (Figure 1), with the biggest increase in temperature observed in November - at what is anyway the hottest time of the year. This is particularly significant for tsetse population dynamics, because our recent analyses indicate not only that this is the time of the year when tsetse mortality is at its highest - but also that the mortality is particularly increased among immature stages and among adults that have just emerged. Further increase in temperature at this time of the year may be expected to result in serious reductions in local tsetse populations.
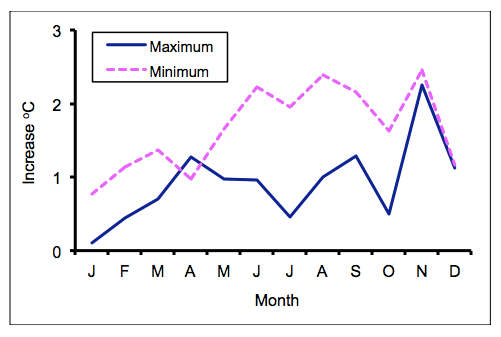
Figure 1. Increase in monthly temperatures from 1959 to 2014. Data from the regression of mean monthly minimum and maximum temperatures against year number.
The ways in which the abundance of tsetse at Rekomitjie has changed since 1959 are being gauged from changes that have occurred in the numbers of tsetse caught from various baits. Such work is complicated since catches are only an indirect measure of tsetse abundance, being dependent on the availability of the population to the sampling system, which varies according to season, the species of tsetse, and the type and location of the sampling device. Nevertheless, it seems that there has been a serious decline in tsetse abundance. For example, catches of G. pallidipes from bait oxen stationed in riverine vegetation in various seasons in 1959/60 were about 20 times greater than the catches that have been made in the last three years by the same method, in the same seasons and situation. Other preliminary data suggest that the past population of G. morsitans was about five times greater than at present.
Types of model
To predict the way that the tsetse population will change in future decades it is necessary to have first a credible model of the way that the population has changed in the past. There are two types of model that could be considered for such work. First, there is the correlation type of model which looks at a wide range of environmental variables, such as temperature, rainfall, settlement, vegetation and land use in different parts of a study area and then attempts to correlate these with the local abundance of tsetse, or sometimes with just the presence or absence of the flies. Extrapolation of the way that these variables have changed in the past can then lead to predictions of future populations. While this may be the simplest and easiest type of predictive modelling, it has the shortcoming that it depends on correlation rather than causation, so that it is a mere “rule of thumb”, even where it can be a sophisticated one. The problem is that the basic causes of changes in population levels are not established in these models, so that there is doubt about the exportability of the model. The danger can be illustrated by reference to the rules of thumb about the weather. For example, the ancient adage:
“Red sky at night, shepherds delight, Red sky at morning shepherds warning”,
relies on correlations that might be valid in the places where it was coined, but it cannot possibly be true at Rekomitjie where for much of the year the sky is red at both morning and night.
Second, there is the type of model that attempts to understand directly the basic causes and effects in the relationships between tsetse and their environment, giving particular attention to environmental effects on birth and death rates of the flies, so establishing the growth rate and hence the extent to which the population can increase or decrease in different circumstances. The more experimental detail we know about the fundamental causes and effects the more confident we can be about predicting the changes in tsetse abundance at any time or in any location. Admittedly, however, the best policy might be to rely on both sorts of modelling, at least in the early stages. For example, the change in tsetse abundance in the last 55 years at Rekomitjie seems to correlate better with temperature rather than with rainfall, so suggesting that the effects of temperature on birth and death rates might be the most useful meteorological aspect of modelling cause and effect.
The seemingly over-riding importance of temperature is hardly surprising since the life-style of the tsetse makes it largely independent of available water. Thus, whereas mosquitoes, horse flies, other biting flies and even house flies require water, or at least some moist environment, where they deposit large numbers of eggs, tsetse have quite different, indeed unique, requirements. Tsetse produce only one egg at roughly 9-day intervals: the egg hatches and is retained in the uterus, where three larval stages are nourished via modified “milk” glands. The pupa that is eventually produced can weigh more than its mother and has all of the energy and raw materials required to produce the adult fly. The pupae are sheltered in the soil and protected against water loss, and the adults obtain plenty of water in their bloodmeals. In these regards the effective modelling of climate change with tsetse is likely to be simpler than with insects such as mosquitoes which are highly dependent on water bodies for much of their life, and so have population dynamics that can vary greatly according to the different rainfall in various seasons and years.
Preliminary modelling
SACEMA is currently developing a cause and effect model for tsetse, based primarily on mean daily temperatures. It is fortunate, therefore, that good experimental data are available for the way in which temperature affects the development of the various life stages of tsetse, so governing the birth rate. Moreover, we also have good data for the effect of temperature on the death rates of the various life stages. Putting these data together, and making credible allowances for the density dependence of tsetse populations, we have simulated the way that the tsetse population at Rekomitjie has changed in the last 55 years and made predictions for the way that it will change in future if temperatures continue to increase at past rates. The salient result is the prediction that tsetse at Rekomitjie could become locally extinct by about 2050 (Figure 2).
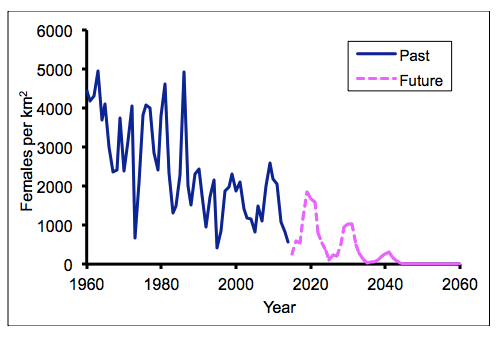
Figure 2. Simulation of past and future abundance of female G. Morsitans per km2 at Rekomitjie. It is assumed that the pattern of rising and falling temperatures of the last decade is serially repeated in each future decade, but with the general level of temperatures rising at previous annual rates.
Further work
The model needs to be refined and validated using more field data from Rekomitjie and Tanzania, and to be expanded to consider spatial effects. For example, while rising temperatures at Rekomitjie might reduce the population there, the flies could transfer to cooler higher ground where extensive human settlement and agriculture occur. This requires the model to be modified to take account of the other great thing that is expected to affect the welfare of tsetse: the type and abundance of hosts. Meanwhile, in anticipation of the possibility of tsetse becoming a greater threat in settled areas, attempts have been made to refine the use of what can be the most appropriate means of tsetse control in agricultural situations, i.e., the restricted application of pyrethroid insecticide to cattle. Particular attention has been given to demonstrating the means of eliminating the risks that pyrethroid contamination of cattle dung can pose to dung beetles - important if the role that such beetles play in soil fertility is to be maintained.
View the original article here http://sacemaquarterly.com/other-infectious-diseas…